原生动物
| 原生动物是原生生物当中较接近动物的一类。简称原虫。它们起初被归类为动物界的一门,为最原始、最简单、最低等的单细胞动物。原生动物是单细胞,细胞内有特化的各种胞器,具有维持生命和延续后代所必需的一切功能,如行动、营养、呼吸、排泄和生殖等。每个原生动物都是一个完整的有机体。有些种类介于植物和动物之间,如眼虫,因为它们能进行光合作用;它们又能运动,并像真正的动物那样进食。原虫很微小,一般只能通过显微镜才能看到。经记录的原生动物约有50000种,其中大约有20000种为化石种。 按照支序分类学说的观点,原生动物是真核生物除去多细胞动物、植物、真菌之外的部分,为并系群,且区分动植物的标准——运动和光合作用均与生物演化分类无关。因此原生动物只是一个集合概念,而不应作为生物分类的单元。原生动物现在被更准确地划分在一个单独的界:原生生物界。 |  1) 草履虫 Paramecium caudatum 2) 团藻 Volvox globator 3) 钟形虫 Vorticella nebulifera 4) 喇叭虫 Stentor polymorphus 5) 变形虫 Amoeba proteus 6) 毛口虫 Trichodina pediculus 7) 单胞藻 Chlamydomonas simplex 8) 唇滴虫 Chilomonas paramecium |
各种原生动物一览
原生动物的伸缩泡和食物泡
生活在淡水的原生动物有1至多个伸缩泡生活在海水中的和寄生的原生动物一般没有伸缩泡。变形虫只有1个伸缩泡,构造简单,位置不定,伸缩泡的周围有小囊泡,线粒体密度较大。草履虫有前后两个伸缩泡,各有1个中央泡和6根辐射伸出的收集管,收集管上有分枝小管与内质网的管道系统相连。伸缩泡的主要功能是调节渗透压。
食物泡根据摄食情况分为两种:含大颗粒食物的吞噬泡和含溶解性营养物质的吞饮泡。食物颗粒或营养物质进入食物泡后,食物泡与细胞质内的溶酶体合并,食物被溶酶体释放的酶消化,未经消化的废物被排出体外。
变形虫
变形虫又音译为“阿米巴”(Amoeba sp.),是一种原生动物,主要生活在清水池塘,或在水流缓慢藻类较多的浅水中,以至一般泥土也可找到,亦可成寄生虫寄生在其他生物里面。变形虫是一种极小的原生动物,全身直径通常只有1%厘米,最大的变形虫直径也只有0.4毫米,用肉眼看,不过是一个模模糊糊的小白点。
变形虫身体仅由一个细胞构成,没有固定的外形,可以任意改变体形。它的细胞膜纤薄;由于原生质的流动,使身体表面生出无定形的指状、叶状或针状的突起,称为“伪足”,身体即借此而移动。身体的形状轮廓也会随伪足的伸缩而有变化。伪足间可自由包围融合,借此包裹事物进行消化。其伪足的主要功能为运动和摄食。它们一般是以单细胞藻类、小型单细胞动物作为食物。自然界常见的为大变形虫,它可吞噬草履虫。当碰到食物时,变形虫会伸出伪足进行包围,由细胞质里面的食物胞消化。变形虫细胞质里面本身有伸缩泡及食物泡,伸缩泡作用是排除变形虫体内过多水分,而食物泡的功能则是消化食物养分。消化好的食物会进入周围的细胞质中;不能消化的物质,就会通过质膜排出体外,称排遗。
变形虫伸出伪足吞食细菌
变形虫与其他生物一样需要利用能量进行呼吸作用。而变形虫的呼吸作用中,所吸入的氧和排出的二氧化碳,都是由细胞膜负责。至于繁殖方式亦相当简单,主要靠有丝分裂繁殖,即原来的遗传物质先复制,然后连同整个细胞一分为二;遗传功能由细胞核负责,跟其他生物一样。
 |
 |
变形虫细胞质的流动
草履虫
草履虫(Paramoecium)属于原生动物门,纤毛虫纲。
草履虫的身体呈圆筒形,前端较圆,中后部较宽,后端较尖,平面看形状像倒置的草底鞋,故名。草履虫体长只有80~300微米,常见的是大草履虫(Paramoecium caudatum),又叫尾草履虫。全身身体表面包着一层膜,膜上密密地长着许多纤毛,靠纤毛的划动在水中旋转运动。身体一侧有一条自前端斜向腰部的凹入小沟,称口沟,沟底有口,相当于草履虫的“嘴巴”。沟内有较为长密的纤毛,口沟内的密长的纤毛摆动时,能把水里的细菌和有机碎屑作为食物摆进口沟,再进入草履虫体内,供其慢慢消化吸收。残渣由一个叫肛点的小孔排出。
草履虫有一大核,一小核,为世界上最小、最简单的动物之一,生活在有淡水的地方,肉眼无法看见,需用复式显微镜观察。大多数单细胞生物可以借由细胞分裂产生两个大小相近的新个体。草履虫靠身体的表膜吸收水里的氧气,排出二氧化碳。
   |
草履虫的分裂
草履虫的细胞器
|
功能
|
口沟
| 取食 |
表膜
| 呼吸(吸收水里的氧气,排出二氧化碳) |
大核
| 营养代谢 |
小核
| 生殖作用 |
食物泡
| 食物泡是草履虫进行胞吞作用产生的,进入细胞后将与初级溶酶体融合形成次级溶酶体。 |
伸缩泡及收集管
| 收集代谢废物和多余的水,并排出体外 |
肛点
| 排出食物残渣 |
纤毛
| 运动 |
草履虫伸缩泡的伸缩
草履虫的进食
眼虫
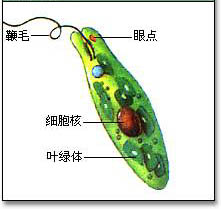
眼虫藻(学名:Euglena sp.)是生物里的一个属,属于裸藻纲。其名字的来源是因为它们有眼斑,它与趋光性有关。眼虫是一种既可定为植物,也可定为动物的单细胞生物。
眼虫为长梭形或圆柱形而带扁平的单细胞藻体,由前端小凹陷生出细长鞭毛一条,其运行方式犹如螺旋桨,其推进能使得眼虫向前运动;鞭毛基部附近有红色小点能感光,称眼点;眼虫没有细胞壁,有一层富有弹性的表膜,所以身体可以伸缩变形,少数种类表膜很硬,不能变形。
它们主要生活在淡水,也有在湿土表面,在含有有机物较多的水中,生长旺盛时,看上去水成为绿色的一片。这种绿色代表了眼虫大多数种有叶绿体,可以进行光合作用(自养型生物)。光对于眼虫来说是性命攸关的。但也有种类的眼虫是异养型生物,它们靠溶在水中的物质生存。如果没有光,它就会分解自身存储的养料或者是摄取周围的养料度日。也有很多种类同时行两种营养方式,属于兼养型生物。
在眼虫的细胞质内有叶绿体(chloroplast)。叶绿体的形状(如卵圆形、盘状、片状、带状、星状等)、大小、数量及其结构(有无蛋白核及副淀粉鞘)为眼虫属、种的分类特征。眼虫主要通过叶绿素在有光的条件下利用光能进行光合作用,把二氧化碳和水合成糖类,这种营养方式(与一般绿色植物相同),称为光合营养(phototrophy)。制造的过多食物形成一些半透明的副淀粉粒(Paramylum granule)储存在细胞质中。副淀粉粒与淀粉相似,是糖类的一种,但与碘作用不呈蓝紫色.副淀粉粒是眼虫类特征之一,其形状大小也是其分类的依据。在无光的条件下,眼虫也可通过体表吸收溶解于水中的有机物质。这种营养方式称为渗透营养(osmotrophy)。
眼虫前端的胞口是否取食固体食物颗粒还有异议。但是已肯定经过胞口可以排出体内过多的水分。在储蓄泡旁边有一个大的伸缩泡(contractile vacuole),它的主要功能是调节水分平衡。液泡根据身处环境的溶液浓度的不同,收缩扩大的速度也不同。收集细胞质中过多的水分(其中也有溶解的代谢废物)。排入储蓄泡,再经胞口排出体外。
当眼虫发育成熟后,它会分裂为两个新细胞,或者和另一个细胞进行生育。在环境不良的条件下。如水池干涸.眼虫体变圆,分泌一种胶质形成包囊,将自己包围起来。刚形成的包囊、可见有眼点,绿色,以后逐流变为黄色,眼点消失,代谢降低,可以生活很久,随风散布于各处。当环境适合时,虫体破囊而出,在出囊前进行一次或几次纵分裂。包囊形成对眼虫度过不良环境是一种很好的适应。
 |  |
眼点的放大观察(1250x)









.jpg)





























{kind=link}